Dong Lab
Research Overview
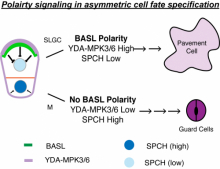
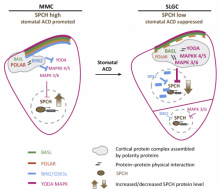
Cell polarity, in both animals and plants, is of paramount importance for many developmental and physiological processes. In the future, my lab will continue to use Arabidopsis as a model system, by studying BASL (Breaking of Asymmetry in the Stomatal Lineage) and other newly identified factors, to investigate how proteins become polarly localized, how polarity proteins are involved in establishment of cellular asymmetry, and how cell polarity is instructive of cell fate and differentiation in plants.
Contact Information
Waksman Institute
190 Frelinghuysen Road
Dong Lab, Office 1012A
Piscataway, NJ 08854
United States
Selected Publications
Complete list of publications: [Google Scholar] [Pubmed]

Current Lab Members
Principal Investigator
-

Dr. Juan Dong
dong@waksman.rutgers.edu

Research Associates
-

Xiaoyu Guo
xyguo@waksman.rutgers.edu

Postdoctoral Researchers
-

Xue Ding
xue.ding@rutgers.edu
-
Aobo Huang
huangaobo@waksman.rutgers.edu
-

Deepanjali Verma
deepanjali@waksman.rutgers.edu
-

Yi Zhang
yi.zhang@waksman.rutgers.edu




Graduate Students
-

Dongmeng Li
dongmeng.li@rutgers.edu
-

Sanqiang Zhang
sz601@waksman.rutgers.edu


Undergraduate Students
-
Shani Gregg
ssg120@scarletmail.rutgers.edu
Alumni
Former Research Associates
- Ying Zhang
Former Postdoctoral Fellows
- Xueyi Xue
Former Graduate Students
- Lu Wang
- Chao Bian
- Wanchen Shao
Former Undergraduate Students
- Gaurav Pathak
- Kavya Mehndiratta